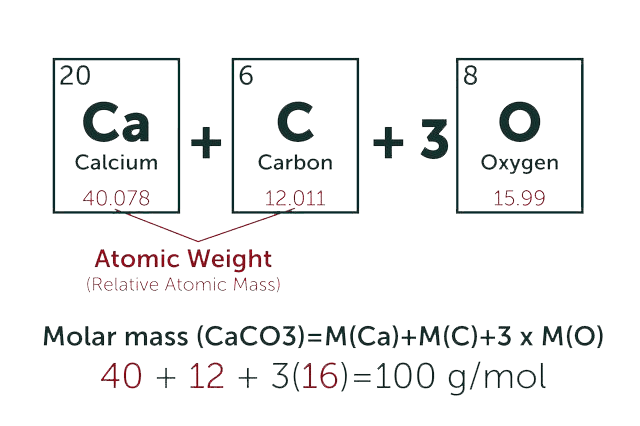
Filipin III
* Please be kindly noted products are not for therapeutic use. We do not sell to patients.

| Category | Antibiotics |
| Catalog number | BBF-01714 |
| CAS | 480-49-9 |
| Molecular Weight | 654.83 |
| Molecular Formula | C35H58O11 |
| Purity | >98% |
Online Inquiry
Description
Filipin III is a polyene macrolide antibiotic produced by Streptomyces filipinensis and Str. durhamensis. It has anti-fungal activity and has the effect of inhibiting Mycoplasma Lebsiella. It also has an effect on the diseases caused by Aspergillus penicillium, stalk mold and other crops such as peas and tomatoes.
Specification
| Synonyms | 14-Deoxylagosin |
| Storage | -20°C |
| IUPAC Name | (3R,4S,6S,8S,10R,12R,14R,16S,17E,19E,21E,23E,25E,27S,28R)-4,6,8,10,12,14,16,27-octahydroxy-3-[(1R)-1-hydroxyhexyl]-17,28-dimethyl-1-oxacyclooctacosa-17,19,21,23,25-pentaen-2-one |
| Canonical SMILES | CCCCCC(C1C(CC(CC(CC(CC(CC(CC(C(=CC=CC=CC=CC=CC(C(OC1=O)C)O)C)O)O)O)O)O)O)O)O |
| InChI | InChI=1S/C35H58O11/c1-4-5-11-16-31(42)34-33(44)22-29(40)20-27(38)18-25(36)17-26(37)19-28(39)21-32(43)23(2)14-12-9-7-6-8-10-13-15-30(41)24(3)46-35(34)45/h6-10,12-15,24-34,36-44H,4-5,11,16-22H2,1-3H3/b7-6+,10-8+,12-9+,15-13+,23-14+/t24-,25+,26-,27+,28-,29+,30+,31-,32+,33+,34-/m1/s1 |
| InChI Key | IMQSIXYSKPIGPD-YQRUMEKGSA-N |
| Source | Streptomyces sp. |
Properties
| Appearance | Pale yellow solid |
| Antibiotic Activity Spectrum | fungi; mycoplasma |
| Boiling Point | 913.9°C at 760 mmHg |
| Density | 1.161 g/cm3 |
| Solubility | Soluble in DMSO |
Reference Reading
1. Solution NMR structures of the polyene macrolide antibiotic filipin III
J Lancelin, L Volpon FEBS Lett . 2000 Jul 28;478(1-2):137-40. doi: 10.1016/s0014-5793(00)01824-x.
The solution structure of filipin III, an antifungal polyene macrolide biosynthesized by Streptomyces filipinensis and widely used for the detection and the quantitation of cholesterol in biomembranes, has been calculated with a set of geometrical restraints derived from 1H NMR in DMSO-d(6) at 25 degrees C. Filipin III appears as a rod-shaped molecule of 18 A length. Its amphiphilic structure is made of an all-syn 1,3-polyol motif, stabilized by intramolecular hydrogen bonds on one side, and a conjugated pentaene moiety on the other side of the molecule. The overall shape is comparable to cholesterol, and the molecular structure of filipin III affords a first molecular basis to the comprehensive understanding of the interactions possible in the filipin III-cholesterol complex which is still unknown at the atomic resolution.
2. Permeabilizing action of filipin III on model membranes through a filipin-phospholipid binding
J Milhaud Biochim Biophys Acta . 1992 Apr 13;1105(2):307-18. doi: 10.1016/0005-2736(92)90209-5.
The binding of the pentaene antibiotic filipin to egg-yolk phosphatidylcholine (EPC) and dimyristoylphosphatidylcholine (DMPC) unilamellar vesicles, has been studied by ultraviolet (UV) absorption and circular dichroism (CD). A stoichiometry of one molecule of filipin for five molecules of phospholipid was demonstrated by CD when phospholipids were in fluid phase. The similarity of the CD spectra with EPC and DMPC established a similar filipin-phospholipid assemblage in both membranes. We therefore postulated that filipin incorporation leads to the formation of gel-like domains in fluid EPC membranes as previously demonstrated for fluid DMPC membranes (Milhaud, J., Mazerski J., Bolard, J. and Dufoure, E.J. (1989) Eur. Biophys. J. 17, 151-158). The release of fluorescent probes (carboxyfluorescein (CF) or calcein (CC)), entrapped in EPC small unilamellar vesicles (SUV), due to the action of filipin, was followed by fluorescence and CD measurements concomitantly. The following observations were made. (1) The percentage of released probe, as a function of the filipin/phospholipid molar ratios, was the same whether or not membranes contained cholesterol. (2) The permeabilization of vesicles proceeded concomitantly with filipin-phospholipid binding while filipin-cholesterol binding leveled off. (3) The release of the content of vesicles occurred by an all-or-none mechanism leaving the depleted vesicles intact. From these observations and from the previous structural findings, a new interpretation of the action of filipin is proposed. Precluding any disruptive effect, inducement of permeability would result from the high intrinsic permeability of the interfacial region at the boundaries of the gel-like domains corresponding to the filipin-phospholipid aggregates. Additionally, we obtained the permeability coefficients for the anionic forms of CC and CF across EPC SUV, 0.6.10(-10) cm s-1 and 2.10(-10) cm s-1, respectively, as compared to 2.5.10(-14) cm s-1 for the counterion Na+ (Hauser, H, Oldani, D. and Phillips, M.C. (1973) Biochemistry 12, 4507-4517).
3. Cholesterol reporter molecules
Gerald Gimpl, Katja Gehrig-Burger Biosci Rep . 2007 Dec;27(6):335-58. doi: 10.1007/s10540-007-9060-1.
Cholesterol is a major constituent of the membranes in most eukaryotic cells where it fulfills multiple functions. Cholesterol regulates the physical state of the phospholipid bilayer, affects the activity of several membrane proteins, and is the precursor for steroid hormones and bile acids. Cholesterol plays a crucial role in the formation of membrane microdomains such as "lipid rafts" and caveolae. However, our current understanding on the membrane organization, intracellular distribution and trafficking of cholesterol is rather poor. This is mainly due to inherent difficulties to label and track this small lipid. In this review, we describe different approaches to detect cholesterol in vitro and in vivo. Cholesterol reporter molecules can be classified in two groups: cholesterol binding molecules and cholesterol analogues. The enzyme cholesterol oxidase is used for the determination of cholesterol in serum and food. Susceptibility to cholesterol oxidase can provide information about localization, transfer kinetics, or transbilayer distribution of cholesterol in membranes and cells. The polyene filipin forms a fluorescent complex with cholesterol and is commonly used to visualize the cellular distribution of free cholesterol. Perfringolysin O, a cholesterol binding cytolysin, selectively recognizes cholesterol-rich structures. Photoreactive cholesterol probes are appropriate tools to analyze or to identify cholesterol binding proteins. Among the fluorescent cholesterol analogues one can distinguish probes with intrinsic fluorescence (e.g., dehydroergosterol) from those possessing an attached fluorophore group. We summarize and critically discuss the features of the different cholesterol reporter molecules with a special focus on recent imaging approaches.
Recommended Products
| BBF-01829 | Deoxynojirimycin | Inquiry |
| BBF-00764 | Cerebroside C | Inquiry |
| BBF-01693 | Doxorubicin EP Impurity A (Daunorubicin) | Inquiry |
| BBF-02614 | Nystatin | Inquiry |
| BBF-03754 | CASTANOSPERMINE | Inquiry |
| BBF-01826 | Deoxymannojirimycin | Inquiry |
Bio Calculators
* Our calculator is based on the following equation:
Concentration (start) x Volume (start) = Concentration (final) x Volume (final)
It is commonly abbreviated as: C1V1 = C2V2
* Total Molecular Weight:
g/mol
Tip: Chemical formula is case sensitive. C22H30N4O √ c22h30n40 ╳